
Сви иЛиве садржаји су медицински прегледани или проверени како би се осигурала што већа тачност.
Имамо стриктне смјернице за набавку и само линкамо на угледне медијске странице, академске истраживачке институције и, кад год је то могуће, медицински прегледане студије. Имајте на уму да су бројеви у заградама ([1], [2], итд.) Везе које се могу кликнути на ове студије.
Ако сматрате да је било који од наших садржаја нетачан, застарио или на неки други начин упитан, одаберите га и притисните Цтрл + Ентер.
Хипоталамус
Медицински стручњак за чланак

Последње прегледано: 07.07.2025
Хипоталамус формира доње делове диенцефалона и учествује у формирању дна треће коморе. Хипоталамус обухвата оптичку хијазму, оптички тракт, сиви туберкулум са левком и мамиларна тела.
Оптичка хијазма (chiasma opticum) је попречни гребен формиран од влакана оптичких живаца (II пар кранијалних живаца), делимично прелазећи на супротну страну (формирајући прегиб). Овај гребен се наставља латерално и постериорно са сваке стране у оптички тракт (tratus opticus). Оптички тракт се налази медијално и постериорно од предње перфориране супстанце, савија се око мождане петељке са латералне стране и завршава се са два корена у субкортикалним визуелним центрима. Већи латерални корен (radix lateralis) прилази латералном геникулалном телу, а тањи медијални корен (radix medialis) иде до горњег коликулуса крова средњег мозга.
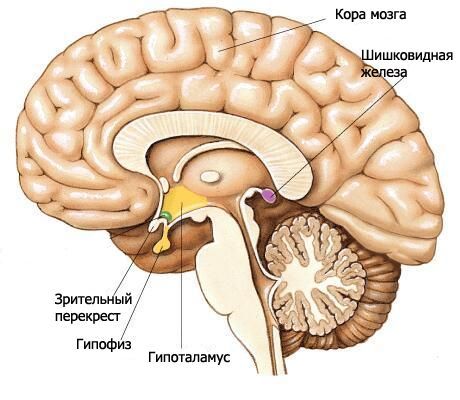
Терминална плоча, која припада теленцефалону, налази се уз предњу површину оптичке хијазме и спаја се са њом. Она затвара предњи део уздужне фисуре великог мозга и састоји се од танког слоја сиве материје, који се у бочним деловима плоче наставља у супстанцу фронталних режњева хемисфера.
Иза оптичке хијазме налази се сиви туберкулус (tuber cinereum), иза којег се налазе мамиларна тела, а са стране су оптички трактови. Испод, сиви туберкулус прелази у левак (infundibulum), који се спаја са хипофизом. Зидове сивог туберкулуса формира танка плоча сиве масе која садржи сиво-туберална језгра (nuclei tuberales). Са стране шупљине треће коморе, сужено удубљење левка се пројектује у предео сивог туберкулуса и даље у левак.
Мамиларна тела (corpora mamillaria) налазе се између сивог туберкула напред и задње перфориране супстанце позади. Изгледају као две мале, пречника око 0,5 цм свака, сферне беле формације. Бела маса се налази само изван мамиларног тела. Унутра је сива маса, у којој се разликују медијално и латерално једро мамиларног тела (nuclei corporis mamillaris mediales et laterales). Стубови форникса се завршавају у мамиларним телима.
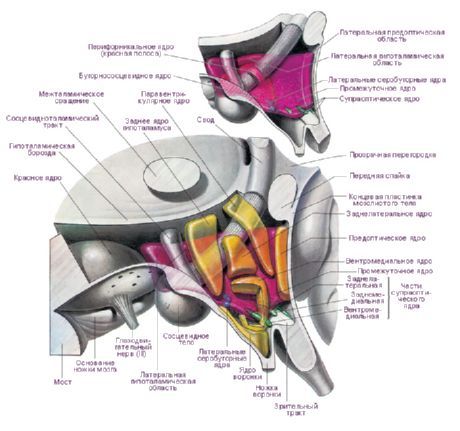
У хипоталамусу постоје три главне хипоталамичке регије - кластери група нервних ћелија различитих облика и величина: предња (regio hypothalamica anterior), средња (regio hypothalamica intermedia) и задња (regio hypothalamica posterior). Кластери нервних ћелија у овим регијама формирају више од 30 језгара хипоталамуса.
Нервне ћелије хипоталамичких једара имају способност да производе секрет (неуросекретију), који се може транспортовати до хипофизе преко наставка истих тих ћелија. Таква једра се називају неуросекреторна једра хипоталамуса. У предњем делу хипоталамуса налазе се супраоптичко (супраоптичко) једро (nucleus supraopticus) и паравентрикуларна једра (nuclei paraventriculares). Наставци ћелија ових једара чине хипоталамуско-хипофизни сноп, који се завршава у задњем режњу хипофизе. Међу групом једара задњег дела хипоталамуса, највећа су медијална и латерална једра мамиларног тела (nuclei corporis mamillaris mediales et laterales) и задње хипоталамично једро (nucleus hypothalamicus posterior). Група језгара интермедијарне хипоталамијске регије обухвата инферомедијална и суперомедијална хипоталамичка језгра (nuclei hypothalamic ventromediales et dorsomediales), дорзално хипоталамично језгро (nucleus hypothalamicus dorsalis), језгро инфундибулариса (nucleus infundibularis), сиво-кртоласта језгра (nuclei tuberales) итд.
Језгра хипоталамуса су повезана прилично сложеним системом аферентних и еферентних путева. Стога, хипоталамус има регулаторни ефекат на бројне вегетативне функције тела. Неуросекреција језгара хипоталамуса може утицати на функције жлезданих ћелија хипофизе, повећавајући или инхибирајући лучење низа хормона, који заузврат регулишу активност других ендокриних жлезда.
Присуство неуралних и хуморалних веза између хипоталамичких језгара и хипофизе омогућило је њихово комбиновање у хипоталамуско-хипофизни систем.
Филогенетске студије су показале да хипоталамус постоји код свих хордата, добро је развијен код водоземаца, а још више код гмизаваца и риба. Птице имају јасно изражену диференцијацију једара. Код сисара је сива маса високо развијена, чије се ћелије диференцирају у једара и поља. Људски хипоталамус се не разликује значајно од хипоталамуса виших сисара.
Постоји велики број класификација хипоталамичких језгара. Е. Грунтел је идентификовао 15 парова језгара, В. Ле Гро Кларк - 16, Х. Куленбек - 29. Најшире коришћена класификација је она В. Ле Гро Кларка. И. Н. Боголепова (1968), на основу горе наведених класификација и узимајући у обзир податке о онтогенези, предлаже поделу хипоталамичких језгара на четири дела:
- предњи, или рострални, пресек (комбинује преоптичку област и предњу групу - В. Ле Гро Кларк) - преоптичке медијалне и латералне области, супрахијазматично једро, супраоптичко једро, паравентрикуларно једро, предња хипоталамичка област;
- средњи медијални део - вентромедијално једро, дорзомедијално једро, инфундибуларно једро, задња хипоталамичка област;
- средњи латерални део - латерална хипоталамичка област, латерално хипоталамично једро, туберолатерално једро, туберомамиларно једро, перифорнично једро;
- задњи, или мамиларни, одељак - медијално мамиларно једро, латерално мамиларно једро.
Анатомске везе хипоталамуса такође разјашњавају његов (функционални) значај. Међу најважнијим аферентним путевима могу се издвојити следећи:
- медијални сноп предњег мозга, чији латерални део повезује хипоталамус са олфакторном сијалицом и туберкулом, периамигдалоидном регијом и хипокампусом, а медијални део са септумом, дијагоналном регијом и каудатним језгром;
- терминална трака, која иде од крајника до предњих делова хипоталамуса;
- влакна која пролазе кроз форникс од хипокампуса до тела мамилара;
- таламо-, стрио- и палидохипоталамске везе;
- из можданог стабла - централни тегментални тракт;
- из мождане коре (орбиталне, темпоралне, паријеталне).
Дакле, водећи извори аферентације су лимбичка формација предњег мозга и ретикуларна формација можданог стабла.
Еферентни системи хипоталамуса такође се могу груписати у три правца:
- силазни системи до ретикуларне формације и кичмене мождине - перивентрикуларни систем влакана који се завршавају у средњем мозгу (лонгитудинални задњи сноп), у аутономним центрима каудалног трупа и кичмене мождине, и мамиларно-тегментални сноп, који иде од мамиларних тела до ретикуларне формације средњег мозга;
- путеви до таламуса од мамиларних тела (маматаламички сноп), који су део затвореног функционалног лимбичког система;
- путеви до хипофизе - хипоталамусно-хипофизни пут од паравентрикуларних (10-20% влакана) и супраоптичких (80-90%) језгара до задњег и делимично средњег режња хипофизе, туберохипофизни пут од вентромедијалних и инфундибуларних језгара до аденохипофизе.
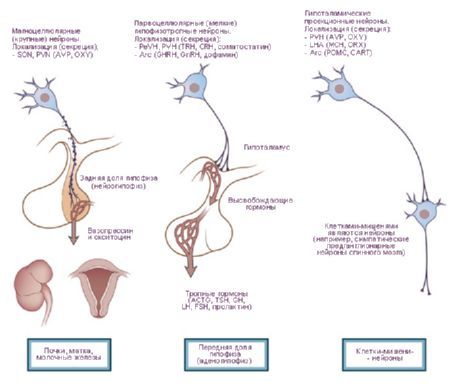
Радови Ј. Рансона (1935) и В. Хеса (1930, 1954, 1968) представили су податке о ширењу и сужавању зенице, повећању и смањењу артеријског притиска, убрзању и успоравању пулса при стимулацији хипоталамуса. На основу ових студија, идентификоване су зоне које врше симпатичке (задњи део хипоталамуса) и парасимпатичке (предњи део) ефекте, а сам хипоталамус је сматран центром који интегрише активност висцералног система који инервира органе и ткива. Међутим, како су се ове студије развијале, откривен је и велики број соматских ефеката, посебно током слободног понашања животиња [Гелхорн Е., 1948]. О. Г. Баклаваџан (1969), при стимулацији различитих делова хипоталамуса, приметио је у неким случајевима реакцију активације у можданој кори, олакшавање моносинаптичких потенцијала кичмене мождине, повећање артеријског притиска, а у другима, супротан ефекат. У овом случају, вегетативне реакције су имале највиши праг. О. Сагер (1962) је открио инхибицију y-система и ЕЕГ синхронизације током дијатермије хипоталамуса, и супротан ефекат током прекомерног загревања. Формира се идеја о хипоталамусу као делу мозга који спроводи интеракцију између регулаторних механизама, интеграцију соматске и вегетативне активности. Са ове тачке гледишта, исправније је поделити хипоталамус не на симпатички и парасимпатички део, већ разликовати динамогене (ерготропне и трофотропне) зоне у њему. Ова класификација је функционалне, биолошке природе и одражава учешће хипоталамуса у спровођењу холистичких бихејвиоралних чинова. Очигледно је да не само вегетативни, већ и соматски систем учествује у одржавању хомеостазе. Ерготропне и трофотропне зоне се налазе у свим деловима хипоталамуса и преклапају се у неким областима. Истовремено, могуће је идентификовати зоне њихове „концентрације“. Дакле, у предњим одељцима (преоптичка зона) трофотропни апарати су јасније заступљени, а у задњим одељцима (мамиларна тела) - ерготропни. Анализа главних аферентних и еферентних веза хипоталамуса са лимбичким и ретикуларним системом баца светло на његову улогу у организацији интегративних облика понашања. Хипоталамус заузима посебан - централни - положај у овом систему како због свог топографског положаја у центру ових формација, тако и као резултат физиолошких карактеристика. Потоње је одређено улогом хипоталамуса као специфично конструисаног дела мозга, посебно осетљивог на промене у унутрашњем окружењу тела, који реагује на најмање флуктуације хуморалних показатеља и формира сврсисходне акте понашања као одговор на те промене.Посебна улога хипоталамуса је предодређена његовом анатомском и функционалном близином хипофизи. Језгра хипоталамуса су подељена на специфична и неспецифична. Прва група обухвата формације које пројектују хипофизу, остатак обухвата друга језгра, чији ефекти стимулације могу варирати у зависности од јачине утицаја. Специфична језгра хипоталамуса имају јасан ефекат и разликују се од других можданих формација по својој способности за неурокринију. То укључује супраоптичка, паравентрикуларна и парвоцелуларна језгра сивог туберкулума. Утврђено је да се антидиуретички хормон (АДХ) формира у супраоптичким и паравентрикуларним језгрима, који се спушта дуж аксона хипоталамо-хипофизног тракта до задњег режња хипофизе. Касније је показано да се у неуронима хипоталамуса формирају фактори ослобађања, који, улазећи у аденохипофизу, регулишу лучење троструких хормона: адренокортикотропног (АЦТХ), лутеинизирајућег (ЛХ), фоликулостимулишућег (ФСХ) и тиреостимулишућег (ТСХ). Зоне формирања имплементационих фактора за АЦТХ и ТСХ су језгра предњег дела медијалне еминенције и преоптичке области, а за ГТГ - задњи делови сивог туберкулума. Утврђено је да хипоталамо-хипофизни снопови код људи садрже око 1 милион нервних влакана.
Несумњиво је да и други делови мозга (медијално-базалне структуре темпоралне регије, ретикуларна формација можданог стабла) учествују у неуроендокриној регулацији. Међутим, најспецифичнији апарат је хипоталамус, који укључује ендокрине жлезде у систем интегралних реакција организма, посебно реакција стресне природе. Трофо- и ерготропни системи располажу не само периферним симпатичким и парасимпатичким системима за обезбеђивање активности, већ и специфичним неурохормоналним апаратима. Хипоталамус-хипофизни систем, који функционише на принципу повратне спреге, у великој мери је саморегулишући. Активност формирања имплементационих фактора такође је одређена нивоом хормона у периферној крви.
Дакле, хипоталамус је важна компонента лимбичког и ретикуларног система мозга, али, будући да је укључен у ове системе, задржава своје специфичне „улазе“ у облику посебне осетљивости на промене у унутрашњем окружењу, као и специфичне „излазе“ кроз хипоталамус-хипофизни систем, паравентрикуларне везе са вегетативним формацијама које леже испод, као и кроз таламус и ретикуларну формацију можданог стабла до кортекса и кичмене мождине.
 [
[ Шта треба испитати?