
Сви иЛиве садржаји су медицински прегледани или проверени како би се осигурала што већа тачност.
Имамо стриктне смјернице за набавку и само линкамо на угледне медијске странице, академске истраживачке институције и, кад год је то могуће, медицински прегледане студије. Имајте на уму да су бројеви у заградама ([1], [2], итд.) Везе које се могу кликнути на ове студије.
Ако сматрате да је било који од наших садржаја нетачан, застарио или на неки други начин упитан, одаберите га и притисните Цтрл + Ентер.
Синтеза, секреција и метаболизам катехоламина
Медицински стручњак за чланак

Последње прегледано: 06.07.2025
Надбубрежна срж производи једињења са структуром која је далеко од стероида. Она садрже 3,4-дихидроксифенил (катехол) језгро и називају се катехоламини. Ту спадају адреналин, норадреналин и допамин бета-хидрокситирамин.
Редослед синтезе катехоламина је прилично једноставан: тирозин → дихидроксифенилаланин (ДОПА) → допамин → норепинефрин → адреналин. Тирозин улази у организам храном, али се може формирати и из фенилаланина у јетри под дејством фенилаланин хидроксилазе. Коначни производи конверзије тирозина у ткивима су различити. У надбубрежној медули процес се одвија до фазе стварања адреналина, у завршецима симпатичких нерава - норепинефрина, у неким неуронима централног нервног система синтеза катехоламина се завршава стварањем допамина.
Конверзију тирозина у ДОПА катализује тирозин хидроксилаза, чији су кофактори тетрахидробиоптерин и кисеоник. Верује се да овај ензим ограничава брзину целог процеса биосинтезе катехоламина и да га инхибирају крајњи производи процеса. Тирозин хидроксилаза је главна мета регулаторних ефеката на биосинтезу катехоламина.
Конверзију ДОПА у допамин катализује ензим ДОПА декарбоксилаза (кофактор - пиридоксал фосфат), који је релативно неспецифичан и декарбоксилише друге ароматичне Л-аминокиселине. Међутим, постоје индикације о могућности модификације синтезе катехоламина променом активности овог ензима. Неким неуронима недостају ензими за даљу конверзију допамина, а он је коначни производ. Друга ткива садрже допамин бета-хидроксилазу (кофактори - бакар, аскорбинска киселина и кисеоник), која претвара допамин у норепинефрин. У надбубрежној медули (али не и у завршецима симпатичких нерава) присутан је фенилетаноламин - метилтрансфераза која формира адреналин из норепинефрина. У овом случају, С-аденозилметионин служи као донор метил група.
Важно је запамтити да синтезу фенилетаноламин-N-метилтрансферазе индукују глукокортикоиди који улазе у срж из кортекса преко порталног венског система. Ово може објаснити чињеницу да су две различите ендокрине жлезде комбиноване у једном органу. Значај глукокортикоида за синтезу адреналина наглашен је чињеницом да се ћелије надбубрежне сржи које производе норадреналин налазе око артеријских крвних судова, док ћелије које производе адреналин примају крв углавном из венских синуса који се налазе у кори надбубрежне жлезде.
Разградња катехоламина се одвија углавном под утицајем два ензимска система: катехол-О-метилтрансферазе (COMT) и моноамин оксидазе (MAO). Главни путеви разградње адреналина и норадреналина су шематски приказани на слици 54. Под утицајем COMT-а у присуству донора метил групе S-адренозилметионина, катехоламини се претварају у норметанефрин и метанефрин (3-O-метил деривати норадреналина и адреналина), који се, под утицајем MAO, претварају у алдехиде, а затим (у присуству алдехид оксидазе) у ванилилманделинску киселину (VMA), главни производ разградње норадреналина и адреналина. У истом случају, када су катехоламини првобитно изложени MAO уместо COMT-а, они се претварају у 3,4-диоксоманделин алдехид, а затим, под утицајем алдехид оксидазе и COMT-а, у 3,4-диоксоманделинску киселину и VMC. У присуству алкохол дехидрогеназе, из катехоламина може да се формира 3-метокси-4-оксифенилгликол, који је главни крајњи производ разградње адреналина и норадреналина у ЦНС-у.
Разградња допамина је слична, осим што његовим метаболитима недостаје хидроксилна група на бета-угљениковом атому, па се уместо ванилилманделне киселине формира хомованилинска киселина (HVA) или 3-метокси-4-хидроксифенилсирћетна киселина.
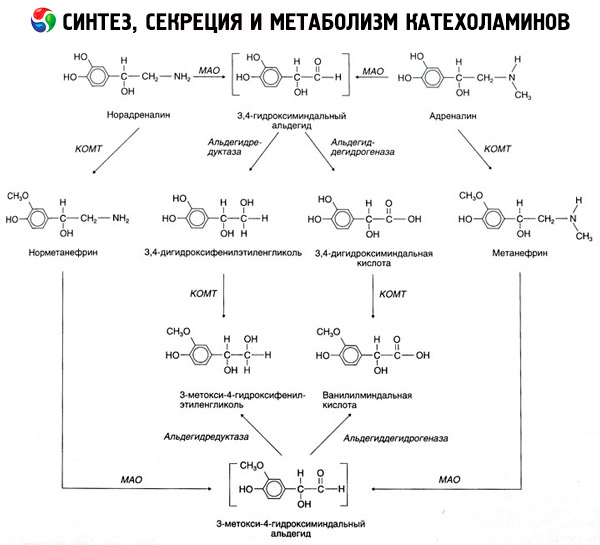
Такође се постулира постојање киноидног пута оксидације молекула катехоламина, који може произвести међупроизводе са израженом биолошком активношћу.
Норепинефрин и адреналин настали под дејством цитосолних ензима у симпатичким нервним завршецима и надбубрежној медули улазе у секреторне грануле, што их штити од дејства ензима за разградњу. Хватање катехоламина гранулама захтева утрошак енергије. У хромафинским гранулама надбубрежне медуле, катехоламини су чврсто везани за АТП (у односу 4:1) и специфичне протеине - хромогранине, што спречава дифузију хормона из гранула у цитоплазму.
Директни стимулус за лучење катехоламина је очигледно продирање калцијума у ћелију, стимулишући егзоцитозу (фузија мембране гранула са површином ћелије и њихово пуцање са потпуним ослобађањем растворљивог садржаја - катехоламина, допамин бета-хидроксилазе, АТП-а и хромогранина - у екстрацелуларну течност).
Физиолошки ефекти катехоламина и њихов механизам деловања
Ефекти катехоламина почињу интеракцијом са специфичним рецепторима циљних ћелија. Док су рецептори за тироидне и стероидне хормоне локализовани унутар ћелија, рецептори за катехоламине (као и ацетилхолин и пептидне хормоне) присутни су на спољашњој површини ћелије.
Одавно је утврђено да су у односу на неке реакције адреналин или норадреналин ефикаснији од синтетичког катехоламина изопротеренола, док је у односу на друге ефекат изопротеренола супериорнији у односу на дејство адреналина или норадреналина. На основу тога развијена је концепција о присуству две врсте адренорецептора у ткивима: алфа и бета, а у неким од њих може бити присутан само један од ова два типа. Изопротеренол је најмоћнији агонист бета-адренорецептора, док је синтетичко једињење фенилефрин најмоћнији агонист алфа-адренорецептора. Природни катехоламини - адреналин и норадреналин - способни су да интерагују са рецепторима оба типа, али адреналин показује већи афинитет за бета-, а норадреналин - за алфа-рецепторе.
Катехоламини активирају срчане бета-адренергичке рецепторе јаче него бета-рецепторе глатких мишића, што је омогућило да се бета тип подели на подтипове: бета1-рецептори (срце, масне ћелије) и бета2-рецептори (бронхије, крвни судови итд.). Дејство изопротеренола на бета1-рецепторе премашује дејство адреналина и норадреналина само 10 пута, док на бета2-рецепторе делује 100-1000 пута јаче од природних катехоламина.
Употреба специфичних антагониста (фентоламин и феноксибензамин за алфа- и пропранолол за бета-рецепторе) потврдила је адекватност класификације адренорецептора. Допамин је способан да интерагује и са алфа- и са бета-рецепторима, али различита ткива (мозак, хипофиза, крвни судови) такође имају своје допаминергичке рецепторе, чији је специфични блокатор халоперидол. Број бета-рецептора варира од 1000 до 2000 по ћелији. Биолошки ефекти катехоламина посредовани бета-рецепторима обично су повезани са активацијом аденилат циклазе и повећањем интрацелуларног садржаја цАМП-а. Иако су рецептор и ензим функционално повезани, то су различити макромолекули. Гванозин трифосфат (ГТП) и други пурински нуклеотиди учествују у модулацији активности аденилат циклазе под утицајем хормон-рецепторског комплекса. Повећавањем активности ензима, изгледа да смањују афинитет бета рецептора за агонисте.
Феномен повећане осетљивости денервисаних структура је одавно познат. Напротив, продужено излагање агонистима смањује осетљивост циљних ткива. Проучавање бета рецептора омогућило је објашњење ових феномена. Показано је да продужено излагање изопротеренолу доводи до губитка осетљивости аденилат циклазе због смањења броја бета рецептора.
Процес десензитизације не захтева активацију синтезе протеина и вероватно је последица постепеног формирања иреверзибилних хормон-рецепторских комплекса. Напротив, уношење 6-оксидопамина, који уништава симпатичке завршетке, прати повећање броја реагујућих бета-рецептора у ткивима. Могуће је да повећање активности симпатичког нервног система такође изазива десензитизацију крвних судова и масног ткива повезану са старењем у односу на катехоламине.
Број адренорецептора у различитим органима може бити контролисан другим хормонима. Тако, естрадиол повећава, а прогестерон смањује број алфа-адренорецептора у материци, што је праћено одговарајућим повећањем и смањењем њеног контрактилног одговора на катехоламине. Ако је интрацелуларни „секундни гласник“ формиран дејством агониста бета-рецептора свакако цАМП, онда је ситуација у погледу преносиоца алфа-адренергичких ефеката компликованија. Претпоставља се постојање различитих механизама: смањење нивоа цАМП-а, повећање садржаја цАМП-а, модулација динамике ћелијског калцијума итд.
Да би се репродуковали различити ефекти у телу, обично су потребне дозе адреналина које су 5-10 пута мање од норадреналина. Иако је овај други ефикаснији у односу на α- и β1-адренорецепторе, важно је запамтити да су оба ендогена катехоламина способна да интерагују и са α- и β-рецепторима. Стога, биолошки одговор датог органа на адренергичку активацију у великој мери зависи од врсте рецептора присутних у њему. Међутим, то не значи да је селективна активација нервне или хуморалне везе симпатичко-адреналног система немогућа. У већини случајева примећује се повећана активност његових различитих веза. Стога је општеприхваћено да хипогликемија рефлексно активира надбубрежну медулу, док је смањење крвног притиска (постурална хипотензија) праћено углавном ослобађањем норадреналина из завршетака симпатичких нерава.
Адренорецептори и ефекти њихове активације у различитим ткивима
Систем, орган |
Тип адренергичког рецептора |
Реакција |
Кардиоваскуларни систем: |
||
Срце |
Бета |
Повећан број откуцаја срца, проводљивост и контрактилност |
Артериоле: |
||
Кожа и слузокоже |
Алфа |
Смањење |
Скелетни мишићи |
Бета |
Експанзија Контракција |
Абдоминални органи |
Алфа (више) |
Смањење |
Бета |
Продужетак |
|
Вене |
Алфа |
Смањење |
Респираторни систем: |
||
Мишићи бронхија |
Бета |
Продужетак |
Систем за варење: |
||
Стомак |
Бета |
Смањене моторичке способности |
Црева |
Алфа |
Контракција сфинктера |
Слезина |
Алфа |
Смањење |
Бета |
Опуштање |
|
Егзокрини панкреас |
Алфа |
Смањена секреција |
Урогенитални систем: |
Алфа |
Контракција сфинктера |
Бешика |
Бета |
Опуштање избацивачког мишића |
Мушки гениталије |
Алфа |
Ејакулација |
Очи |
Алфа |
Проширење зеница |
Кожа |
Алфа |
Повећано знојење |
Пљувачне жлезде |
Алфа |
Излучивање калијума и воде |
Бета |
Секреција амилазе |
|
Ендокрине жлезде: |
||
Острвца панкреаса |
||
Бета ћелије |
Алфа (више) |
Смањена секреција инсулина |
Бета |
Повећана секреција инсулина |
|
Алфа ћелије |
Бета |
Повећана секреција глукагона |
8-ћелијска |
Бета |
Повећана секреција соматостатина |
Хипоталамус и хипофиза: |
||
Соматотрофи |
Алфа |
Повећана секреција СТХ |
Бета |
Смањена секреција СТХ |
|
Лактотрофи |
Алфа |
Смањена секреција пролактина |
Тиротрофи |
Алфа |
Смањена секреција ТСХ |
Кортикотрофи |
Алфа |
Повећана секреција АЦТХ |
| бета | Смањена секреција АЦТХ | |
Штитна жлезда: |
||
Фоликуларне ћелије |
Алфа |
Смањена секреција тироксина |
Бета |
Повећана секреција тироксина |
|
Парафоликуларне (К) ћелије |
Бета |
Повећана секреција калцитонина |
Паратироидне жлезде |
Бета |
Повећана секреција ПТХ |
Бубрези |
Бета |
Повећана секреција ренина |
Стомак |
Бета |
Повећана секреција гастрина |
БX |
Бета |
Повећана потрошња кисеоника |
Јетра |
? |
Повећана гликогенолиза и глуконеогенеза са ослобађањем глукозе; повећана кетогенеза са ослобађањем кетонских тела |
Масно ткиво |
Бета |
Повећана липолиза са ослобађањем слободних масних киселина и глицерола |
Скелетни мишићи |
Бета |
Повећана гликолиза са ослобађањем пирувата и лактата; смањена протеолиза са смањењем ослобађања аланина, глутамина |
Важно је узети у обзир да резултати интравенске примене катехоламина не одражавају увек адекватно ефекте ендогених једињења. Ово се углавном односи на норепинефрин, јер се у телу ослобађа углавном не у крв, већ директно у синаптичке пукотине. Стога, ендогени норепинефрин активира, на пример, не само васкуларне алфа рецепторе (повишен крвни притисак), већ и бета рецепторе срца (убрзан рад срца), док уношење норепинефрина споља доводи углавном до активације васкуларних алфа рецептора и рефлексног (преко вагуса) успоравања срчаног ритма.
Ниске дозе адреналина активирају углавном бета рецепторе мишићних крвних судова и срца, што резултира смањењем периферног васкуларног отпора и повећањем срчаног излаза. У неким случајевима, први ефекат може превладати, а хипотензија се развија након примене адреналина. У вишим дозама, адреналин такође активира алфа рецепторе, што је праћено повећањем периферног васкуларног отпора и, на позадини повећања срчаног излаза, доводи до повећања артеријског притиска. Међутим, његов ефекат на васкуларне бета рецепторе је такође очуван. Као резултат тога, повећање систолног притиска превазилази сличан показатељ дијастолног притиска (повећање пулсног притиска). Увођењем још већих доза, алфа-миметички ефекти адреналина почињу да преовлађују: систолни и дијастолни притисак се повећавају паралелно, као под утицајем норепинефрина.
Утицај катехоламина на метаболизам састоји се од њихових директних и индиректних ефеката. Први се остварују углавном преко бета рецептора. Сложенији процеси су повезани са јетром. Иако се повећана гликогенолиза јетре традиционално сматра резултатом активације бета рецептора, постоје и докази о учешћу алфа рецептора. Индиректни ефекти катехоламина повезани су са модулацијом секреције многих других хормона, као што је инсулин. У дејству адреналина на његову секрецију, алфа адренергичка компонента јасно преовлађује, будући да је показано да је сваки стрес праћен инхибицијом секреције инсулина.
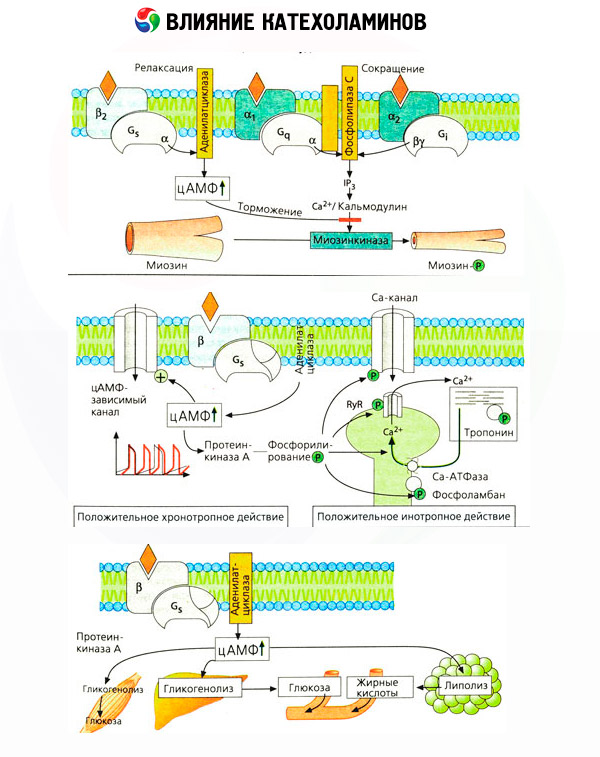
Комбинација директних и индиректних ефеката катехоламина узрокује хипергликемију, повезану не само са повећаном производњом глукозе у јетри, већ и са инхибицијом њеног искоришћавања од стране периферних ткива. Убрзање липолизу узрокује хиперлипацидемију са повећаном испоруком масних киселина у јетру и интензивирањем производње кетонских тела. Повећана гликолиза у мишићима доводи до повећања ослобађања лактата и пирувата у крв, који, заједно са глицеролом ослобођеним из масног ткива, служе као прекурсори глуконеогенезе у јетри.
Регулација секреције катехоламина. Сличност производа и метода реакције симпатичког нервног система и надбубрежне медуле била је основа за обједињавање ових структура у јединствени симпатичко-адренални систем тела са издвајањем његових нервних и хормонских веза. Различити аферентни сигнали су концентрисани у хипоталамусу и центрима кичмене мождине и продужене мождине, одакле потичу еферентне поруке, прелазећи на ћелијска тела преганглионских неурона који се налазе у бочним роговима кичмене мождине на нивоу VIII цервикалног - II-III лумбалног сегмента.
Преганглионски аксони ових ћелија напуштају кичмену мождину и формирају синаптичке везе са неуронима који се налазе у ганглијама симпатичког ланца или са ћелијама надбубрежне медуле. Ова преганглионска влакна су холинергичка. Прва фундаментална разлика између симпатичких постганглионских неурона и хромафинских ћелија надбубрежне медуле је у томе што ове друге преносе холинергички сигнал који примају не нервном проводљивошћу (постганглионски адренергички нерви), већ хуморалним путем, ослобађајући адренергичка једињења у крв. Друга разлика је у томе што постганглионски нерви производе норепинефрин, док ћелије надбубрежне медуле производе углавном адреналин. Ове две супстанце имају различите ефекте на ткива.