
Сви иЛиве садржаји су медицински прегледани или проверени како би се осигурала што већа тачност.
Имамо стриктне смјернице за набавку и само линкамо на угледне медијске странице, академске истраживачке институције и, кад год је то могуће, медицински прегледане студије. Имајте на уму да су бројеви у заградама ([1], [2], итд.) Везе које се могу кликнути на ове студије.
Ако сматрате да је било који од наших садржаја нетачан, застарио или на неки други начин упитан, одаберите га и притисните Цтрл + Ентер.
Јајник
Медицински стручњак за чланак

Последње прегледано: 04.07.2025

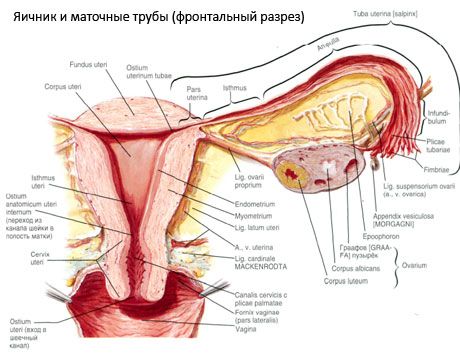
Јајник (овариум; грч. oophoron) је парни орган, женска репродуктивна жлезда, која се налази у карличној дупљи иза широког лигамента материце. У јајницима се развијају и сазревају женске репродуктивне ћелије (јајне ћелије), а формирају се и женски полни хормони који улазе у крв и лимфу. Јајник има јајолики облик, донекле спљоштено у антеропостериорном смеру. Боја јајника је ружичаста. На површини јајника жене која је родила видљива су удубљења и ожиљци - трагови овулације и трансформације жутог тела. Јајник тежи 5-8 г. Димензије јајника су: дужина 2,5-5,5 цм, ширина 1,5-3,0 цм, дебљина - до 2 цм. Јајник има две слободне површине: медијалну површину (facies medialis), окренуту ка карличној дупљи, делимично покривену јајоводом, и бочну површину (facies lateralis), која се наслања на бочни зид карлице, до благо израженог удубљења - јајничке јаме. Ова фоса се налази у углу између спољашњих илијачних крвних судова прекривених перитонеумом на врху и утериних и обтураторних артерија на дну. Иза јајника, уретер одговарајуће стране пролази ретроперитонеално одозго надоле.
Површине јајника прелазе у конвексну слободну (задњу) ивицу (margo liber), напред - у мезентеричну ивицу (margo mesovaricus), причвршћену помоћу кратког набора перитонеума (мезентерија јајника) за задњи листић широког лигамента материце. На овој предњој ивици органа налази се жлебљено удубљење - хилум јајника (hilum ovarii), кроз који артерија и живци улазе у јајник, а излазе вене и лимфни судови. Јајник такође има два краја: заобљени горњи цевасти крај (extremitas tubaria), окренут ка јајоводу, и доњи матерични крај (extremitas utenna), повезан са материцом јајниковим лигаментом (lig. ovarii proprium). Овај лигамент, у облику округлог ужета дебљине око 6 mm, иде од материчног краја јајника до бочног угла материце, који се налази између два листа широког лигамента материце. Лигаментни апарат јајника такође обухвата лигамент суспензоријум јајника (lig.suspensorium ovarii), који представља набор перитонеума који се протеже од зида мале карлице до јајника и садржи крвне судове јајника и снопове влакана унутра. Јајник је фиксиран кратким мезентеријумом (mesovarium), који представља дупликат перитонеума и протеже се од задњег листића широког лигамента материце до мезентеричне ивице јајника. Сами јајници нису покривени перитонеумом. Највећи јајнички руб јајовода је причвршћен за цевасти крај јајника. Топографија јајника зависи од положаја материце, њене величине (током трудноће). Јајници су веома покретни органи мале карличне дупље.
 [
[ Крвни судови и живци јајника
Снабдевање крвљу јајника обезбеђују аа. ет вв. оварицае ет утеринае. Обе јајничке артерије (аа. оварицае дестра ет синистра) потичу са предње површине аорте, одмах испод бубрежних артерија; десна обично потиче из аорте, а лева из бубрежне артерије. Усмеравајући се надоле и латерално дуж предње површине великог псоас мишића, свака јајничка артерија прелази уретер напред (дајући гране ка њему), спољашње илијачне крвне судове, граничну линију и улази у карличну дупљу, где се налази у суспензорном лигаменту јајника. Пратећи у медијалном смеру, јајничка артерија пролази између листова широког лигамента материце испод јајовода, дајући гране ка њему, а затим у мезентеријум јајника; улази у хилум јајника.
Гране оваријалне артерије широко анастомозирају са оваријалним гранама материчне артерије. Венски одлив из јајника се спроводи првенствено у оваријални венски плексус, који се налази у пределу оваријалног хилума. Одавде, одлив крви иде у два смера: кроз материчне и оваријалне вене. Десна оваријална вена има залиске и улива се у доњу шупљу вену. Лева оваријална вена се улива у леву бубрежну вену, иако нема залиске.
Лимфна дренажа из јајника се одвија кроз лимфне судове, посебно обилно у пределу органске капије, где се разликује субоваријални лимфни плексус. Затим се лимфа испушта дуж оваријалних лимфних судова до парааорталних лимфних чворова.
Инервација јајника
Симпатички - обезбеђују га постганглијска влакна из целијачног (соларног), горњег мезентеричног и хипогастричног плексуса; парасимпатички - висцерални сакрални живци.
Структура јајника
Површина јајника је прекривена једнослојним герминативним епителом. Испод њега лежи густи протеински омотач везивног ткива (tunica albuginea). Везивно ткиво јајника формира његову строму (stroma ovarii), богату еластичним влакнима. Супстанца јајника, његов паренхим, подељена је на спољашњи и унутрашњи слој. Унутрашњи слој, који се налази у центру јајника, ближе његовој капији, назива се срж (medulla ovarii). У овом слоју, у растреситом везивном ткиву, налазе се бројни крвни и лимфни судови и живци. Спољашњи слој јајника - кортекс (cortex ovarii) је гушћи. Садржи доста везивног ткива, у коме се налазе сазревајући примарни фоликули јајника (folliculi ovarici primarii), секундарни (везикуларни) фоликули (folliculi ovarici secundarii, s.vesiculosi), као и зрели фоликули, Графови фоликули (folliculi ovarici maturis), као и жута и атретична тела.
Сваки фоликул садржи женску репродуктивну јајну ћелију, или ооцит (овоцитус). Јајна ћелија је пречника до 150 µм, округлог је облика, садржи једро, велику количину цитоплазме, која, поред ћелијских органела, садржи протеинско-липидне инклузије (жуманце), гликоген, неопходан за исхрану јајне ћелије. Јајна ћелија обично потроши своје залихе хранљивих материја у року од 12-24 сата након овулације. Ако не дође до оплодње, јајна ћелија умире.
Људска јајна ћелија има две мембране које је покривају. Унутра се налази цитолема, што је цитоплазматска мембрана јајне ћелије. Споља од цитолеме налази се слој такозваних фоликуларних ћелија, које штите јајну ћелију и обављају функцију стварања хормона - луче естрогене.
Физиолошки положај материце, јајовода и јајника обезбеђују суспензиони, фиксирајући и потпорни апарати који обједињују перитонеум, лигаменте и карлично ткиво. Суспензиони апарат је представљен упареним формацијама, обухвата округле и широке лигаменте материце, праве лигаменте и суспензионе лигаменте јајника. Широки лигаменти материце, прави и суспензиони лигаменти јајника држе материцу у средњем положају. Округли лигаменти вуку фундус материце напред и обезбеђују његов физиолошки нагиб.
Фиксни (сидрени) апарат обезбеђује положај материце у центру мале карлице и практично онемогућава њено померање у стране, напред-назад. Али пошто се лигаментни апарат одваја од материце у њеном доњем делу, могући су нагиби материце у различитим правцима. Фиксни апарат обухвата лигаменте који се налазе у растреситом ткиву карлице и протежу се од доњег дела материце до бочних, предњих и задњих зидова карлице: сакро-магични, кардинални, утеровезикални и везикопубични лигаменти.
Поред мезоварија, разликују се следећи лигаменти јајника:
- суспензорни лигамент јајника, раније означен као инфундибулопелвични лигамент. То је набор перитонеума са крвним судовима (a. et v. ovarica) и лимфним судовима и живцима јајника који пролазе кроз њега, растегнут између бочног зида карлице, лумбалне фасције (у пределу поделе заједничке илијачне артерије на спољашњу и унутрашњу) и горњег (тубалног) краја јајника;
- Прави лигамент јајника пролази између листова широког утерусног лигамента, ближе задњем листу, и повезује доњи крај јајника са бочном ивицом материце. Прави лигамент јајника је причвршћен за материцу између почетка јајовода и округлог лигамента, иза и изнад овог другог. У дебљини лигамента пролазе rr. оварија, који су завршне гране утерусне артерије;
- Апендикуларно-јајни лигамент Лигамент се протеже од мезентерија слепог црева до десног јајника или широког лигамента материце у облику набора перитонеума. Лигамент је непостојан и примећује се код 1/2 - 1/3 жена.
Потпорни апарат је представљен мишићима и фасцијом карличног дна, подељеним на доњи, средњи и горњи (унутрашњи) слој.
Најмоћнији је горњи (унутрашњи) слој мишића, представљен парним мишићем који подиже анус. Састоји се од мишићних снопова који се шире од тртичне кости до карличних костију у три смера (пубокоцигеус, илиокоцигеус и ишиокоцигеус). Овај слој мишића се назива и карлична дијафрагма.
Средњи слој мишића налази се између симфизе, пубичних и исхијалних костију. Средњи слој мишића - урогенитална дијафрагма - заузима предњу половину карличног излаза, кроз који пролазе уретра и вагина. У предњем делу између њених листова налазе се мишићни снопови који формирају спољашњи сфинктер уретре, у задњем делу су мишићни снопови који иду у попречном смеру - дубоки попречни мишић међице.
Доњи (спољашњи) слој мишића карличног дна састоји се од површинских мишића, чији облик подсећа на број 8. То укључује булбокавернозни, ишиокавернозни, спољашњи анални сфинктер и површински попречни перинеални мишић.
Онтогенеза јајника
Процес раста и атрезије фоликула почиње у 20. недељи трудноће, а до тренутка порођаја, у јајницима девојчице остаје до 2 милиона ооцита. До менархе, њихов број се смањује на 300 хиљада. Током целог периода репродуктивног живота, не више од 500 фоликула достиже зрелост и овулира. Почетни раст фоликула не зависи од стимулације ФСХ, ограничен је, а атрезија се брзо јавља. Верује се да су уместо стероидних хормона, локални аутокрини/паракрини пептиди главни регулатор раста и атрезије примарних фоликула. Верује се да процес раста и атрезије фоликула није прекинут никаквим физиолошким процесима. Овај процес се наставља у свим узрастима, укључујући интраутерини период и менопаузу, а прекида га трудноћа, овулација и ановулација. Механизам који покреће раст фоликула и њихов број у сваком специфичном циклусу још увек није јасан.
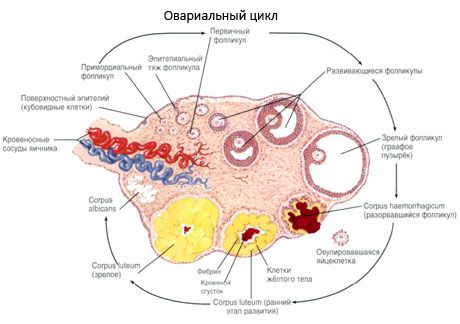
Током свог развоја, фоликул пролази кроз неколико фаза развоја. Примордијалне клице потичу из ендодерма жуманчане кесе, алантоиса и мигрирају у генитални део ембриона у 5-6 недељи трудноће. Као резултат брзе митотске деобе, која се наставља од 6-8 недеље до 16-20 недеље трудноће, у јајницима ембриона се формира до 6-7 милиона ооцита, окружених танким слојем гранулозних ћелија.
Преантрални фоликул - ооцит је окружен мембраном (Zona pellucida). Гранулоза ћелије које окружују ооцит почињу да се размножавају, њихов раст зависи од гонадотропина и корелира са нивоом естрогена. Гранулоза ћелије су мета за ФСХ. У фази преантралног фоликула, гранулозне ћелије су способне да синтетишу три класе стероида: претежно индукује активност ароматазе, главног ензима који претвара андрогене у естрадиол. Верује се да је естрадиол способан да повећа број сопствених рецептора, вршећи директан митогени ефекат на гранулозне ћелије независно од ФСХ. Сматра се паракриним фактором који појачава ефекте ФСХ, укључујући активацију процеса ароматизације.
ФСХ рецептори се појављују на мембранама гранулозних ћелија чим почне раст фоликула. Смањење или повећање ФСХ доводи до промене броја његових рецептора. Ово дејство ФСХ модулирају фактори раста. ФСХ делује преко Г-протеина, система аденилат циклазе, иако је стероидогенеза у фоликулу углавном регулисана ФСХ, у овај процес је укључено много фактора: јонски канали, тирозин киназни рецептори, фосфолипазни систем секундарних гласника.
Улога андрогена у раном развоју фоликула је сложена. Гранулоза ћелије имају андрогене рецепторе. Они нису само супстрат за ароматизацију естрогена изазвану ФСХ-ом, већ могу и појачати ароматизацију при ниским концентрацијама. Када се нивои андрогена повећају, преантралне гранулозне ћелије преференцијално бирају не пут ароматизације естрогена, већ једноставнији пут андрогена преко 5а-редуктазе да би се претвориле у андроген који се не може претворити у естроген, чиме инхибирају активност ароматазе. Овај процес такође инхибира формирање ФСХ и ЛХ рецептора, чиме се зауставља развој фоликула.
Процесом ароматизације, фоликул са високим нивоом андрогена пролази кроз процесе атрезије. Раст и развој фоликула зависи од његове способности да претвори андрогене у естрогене.
У присуству ФСХ, доминантна супстанца фоликуларне течности биће естрогени. У одсуству ФСХ - андрогени. ЛХ је нормално одсутан у фоликуларној течности до средине циклуса. Чим се ниво ЛХ повећа, митотска активност гранулозних ћелија се смањује, јављају се дегенеративне промене и ниво андрогена у фоликулу се повећава. Ниво стероида у фоликуларној течности је виши него у плазми и одражава функционалну активност ћелија јајника: гранулозних и тека ћелија. Ако је једина мета за ФСХ гранулозне ћелије, онда ЛХ има много мета - тека ћелије, стромалне и лутеинске ћелије и гранулозне ћелије. И гранулозне и тека ћелије имају способност стероидогенезе, али у гранулозним ћелијама преовладава ароматазна активност.
Као одговор на ЛХ, тека ћелије производе андрогене, које затим гранулозне ћелије претварају у естрогене путем ароматизације индуковане ФСХ.
Како фоликул расте, тека ћелије почињу да експресују гене за ЛХ рецепторе, П450 сек и 3бета-хидроксистероид дехидрогеназу, инсулину сличан фактор раста (ИГФ-1) синергистички са ЛХ повећава експресију гена, али не стимулише стероидогенезу.
Оваријална стероидогенеза је увек зависна од ЛХ. Како фоликул расте, тека ћелије експресују ензим П450ц17, који формира андроген из холестерола. Гранулоза ћелије немају овај ензим и зависе од тека ћелија да би производиле естрогене из андрогена. За разлику од стероидогенезе, фоликулогенеза је зависна од ФСХ. Како фоликул расте и нивои естрогена се повећавају, активира се механизам повратне спреге - производња ФСХ је инхибирана, што заузврат доводи до смањења активности ароматазе фоликула и, на крају, до фоликуларне атрезије путем апоптозе (програмиране ћелијске смрти).
Механизам повратне спреге естрогена и ФСХ инхибира развој фоликула који су почели да расту, али не и доминантног фоликула. Доминантни фоликул садржи више ФСХ рецептора, који подржавају пролиферацију гранулозних ћелија и ароматизацију андрогена у естрогене. Поред тога, паракрини и аутокрини путеви делују као важан координатор развоја антралног фоликула.
Аутокрини/паракрини регулатор се састоји од пептида (инхибин, активин, фолистатин), које синтетишу гранулозне ћелије као одговор на ФСХ и улазе у фоликуларну течност. Инхибин смањује секрецију ФСХ; активин стимулише ослобађање ФСХ из хипофизе и појачава дејство ФСХ у јајнику; фолистатин сузбија активност ФСХ, могуће везивањем за активин. Након овулације и развоја жутог тела, инхибин је под контролом ЛХ.
На раст и диференцијацију ћелија јајника утичу инсулину слични фактори раста (IGE). IGF-1 делује на гранулозне ћелије, узрокујући повећање цикличног аденозин монофосфата (цАМП), прогестерона, окситоцина, протеогликана и инхибина.
IGF-1 делује на тека ћелије, узрокујући повећану производњу андрогена. Тека ћелије, заузврат, производе фактор туморске некрозе (TNF) и епидермални фактор раста (EGF), које такође регулише FSH.
EGF стимулише пролиферацију гранулозних ћелија. IGF-2 је главни фактор раста у фоликуларној течности, а у њој се налазе и IGF-1, TNF-α, TNF-3 и EGF.
Поремећај паракрине и/или аутокрине регулације функције јајника изгледа да игра улогу у поремећајима овулације и у развоју полицистичних јајника.
Како антрални фоликул расте, садржај естрогена у фоликуларној течности се повећава. На врхунцу њиховог повећања, рецептори за ЛХ се појављују на гранулозним ћелијама, долази до лутеинизације гранулозних ћелија и повећава се производња прогестерона. Дакле, у преовулаторном периоду, повећање производње естрогена узрокује појаву ЛХ рецептора, ЛХ, заузврат, узрокује лутеинизацију гранулозних ћелија и производњу прогестерона. Повећање прогестерона смањује ниво естрогена, што очигледно узрокује други врх ФСХ у средини циклуса.
Сматра се да се овулација јавља 10-12 сати након врхунца ЛХ и 24-36 сати након врхунца естрадиола. Сматра се да ЛХ стимулише редукциону деобу ооцита, лутеинизацију гранулозних ћелија и синтезу прогестерона и простагландина у фоликулу.
Прогестерон појачава активност протеолитичких ензима, који заједно са простагландином учествују у пуцању зида фоликула. Врх ФСХ изазван прогестероном омогућава ооциту да изађе из фоликула претварањем плазминогена у протеолитички ензим плазмин и обезбеђује довољан број ЛХ рецептора за нормалан развој лутеалне фазе.
У року од 3 дана након овулације, гранулозне ћелије се повећавају у величини, а у њима се појављују карактеристичне вакуоле испуњене пигментом, лутеином. Тека-лутеалне ћелије се диференцирају од теке и строме и постају део жутог тела. Капилари који продиру у жуто тело се веома брзо развијају под утицајем фактора ангиогенезе, а са побољшаном васкуларизацијом се повећава производња прогестерона и естрогена. Активност стероидогенезе и животни век жутог тела одређени су нивоом ЛХ. Жуто тело није хомогена ћелијска формација. Поред 2 врсте лутеалних ћелија, садржи ендотелне ћелије, макрофаге, фибробласте итд. Велике лутеалне ћелије производе пептиде (релаксин, окситоцин) и активније су у стероидогенези са већом ароматазном активношћу и већом синтезом прогестерона од малих ћелија.
Врхунац прогестерона јавља се 8. дана након врха ЛХ. Примећено је да се прогестерон и естрадиол епизодно луче у лутеалној фази у корелацији са пулсирајућом лучењем ЛХ. Са формирањем жутог тела, контрола производње инхибина прелази са ФСХ на ЛХ. Инхибин се повећава са повећањем естрадиола пре врха ЛХ и наставља да расте након врха ЛХ, иако нивои естрогена опадају. Иако инхибин и естрадиол луче гранулозне ћелије, они су регулисани различитим путевима. Пад инхибина на крају лутеалне фазе доприноси повећању ФСХ за следећи циклус.
Жуто тело се веома брзо смањује - 9.-11. дана након овулације.
Механизам дегенерације није јасан и није повезан са лутеолитичком улогом естрогена или са механизмом повезаним са рецепторима, као што је примећено у ендометријуму. Постоји још једно објашњење за улогу естрогена које производи жуто тело. Познато је да су естрогени неопходни за синтезу прогестеронских рецептора у ендометријуму. Естрогени лутеалне фазе су вероватно неопходни за промене повезане са прогестероном у ендометријуму након овулације. Неадекватан развој прогестеронских рецептора, као последица неадекватног нивоа естрогена, вероватно је додатни механизам за неплодност и ране губитке трудноће, још један облик недостатка лутеалне фазе. Верује се да се животни век жутог тела успоставља у време овулације. И сигурно ће регресовати ако га не подржава хумани хорионски гонадотропин због трудноће. Дакле, регресија жутог тела доводи до смањења нивоа естрадиола, прогестерона и инхибина. Смањење инхибина уклања његов супресивни ефекат на ФСХ; Смањење естрадиола и прогестерона омогућава веома брзо обнављање секреције GnRH и уклањање механизма повратне спреге из хипофизе. Смањење инхибина и естрадиола, заједно са повећањем GnRH, доводи до пораста FSH у односу на LH. Повећање FSH доводи до раста фоликула са накнадном селекцијом доминантног фоликула, а нови циклус почиње ако не дође до трудноће. Стероидни хормони играју водећу улогу у репродуктивној биологији и општој физиологији. Они одређују људски фенотип, утичу на кардиоваскуларни систем, метаболизам костију, кожу, опште благостање и играју кључну улогу у трудноћи. Дејство стероидних хормона одражава интрацелуларне и генетске механизме који су неопходни за пренос екстрацелуларног сигнала до ћелијског једра како би се изазвао физиолошки одговор.
Естрогени дифузно продиру кроз ћелијску мембрану и везују се за рецепторе који се налазе у ћелијском једру. Комплекс рецептор-стероид се затим везује за ДНК. У циљним ћелијама, ове интеракције доводе до експресије гена, синтезе протеина и специфичне функције ћелија и ткива.
Использованная литература