
Сви иЛиве садржаји су медицински прегледани или проверени како би се осигурала што већа тачност.
Имамо стриктне смјернице за набавку и само линкамо на угледне медијске странице, академске истраживачке институције и, кад год је то могуће, медицински прегледане студије. Имајте на уму да су бројеви у заградама ([1], [2], итд.) Везе које се могу кликнути на ове студије.
Ако сматрате да је било који од наших садржаја нетачан, застарио или на неки други начин упитан, одаберите га и притисните Цтрл + Ентер.
Колера вибрио
Медицински стручњак за чланак

Последње прегледано: 04.07.2025
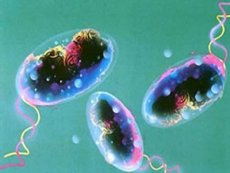
Према СЗО, колера је заразна болест коју карактерише акутна, тешка, дехидрирајућа дијареја са столицом у облику пиринчане воде, што је последица инфекције са Vibrio cholerae. Због чињенице да се карактерише израженом способношћу широког ширења у епидемијама, тешким током и високом стопом смртности, колера се сматра посебно опасном инфекцијом.
Историјска домовина колере је Индија, тачније, делта река Ганг и Брамапутра (данас Источна Индија и Бангладеш), где постоји од памтивека (епидемије колере у овом региону примећене су још 500. године пре нове ере). Дуго постојање ендемског жаришта колере овде објашњава се многим разлозима. Вибрион колере не само да може дуго да преживи у води, већ се и размножава у њој под повољним условима - температурама изнад 12 °C, присуством органске материје. Сви ови услови су евидентни у Индији: тропска клима (просечна годишња температура од 25 до 29 °C), обилне падавине и мочваре, велика густина насељености, посебно у делти реке Ганг, велика количина органске материје у води, континуирано загађење воде канализацијом и изметом током целе године, низак материјални стандард живота и јединствени верски и култни обреди становништва.
У историји епидемија колере могу се разликовати четири периода.
Период I - до 1817. године, када је колера била концентрисана само у Источној и Јужној Азији, углавном у Индији, и није се ширила ван њених граница.
II период - од 1817. до 1926. године. Успостављањем широких економских и других веза између Индије и европских и других земаља, колера је превазишла Индију и, ширећи се дуж путева економских и верских веза, изазвала је 6 пандемија које су однеле милионе људских живота. Русија је била прва од европских земаља у коју је колера продрла. Од 1823. до 1926. године Русија је доживела 57 колерних година. Током овог времена, више од 5,6 милиона људи је оболело од колере, а 2,14 милиона људи је умрло од ње („40%).
III период - од 1926. до 1961. године Колера се вратила у своје главно ендемско жариште, и почео је период релативног благостања. Чинило се да ће развојем савремених система за пречишћавање воде за пиће, уклањање и дезинфекцију отпадних вода и развојем посебних мера против колере, укључујући и стварање карантинске службе, земље света бити поуздано заштићене од још једне инвазије колере.
Четврти период је почео 1961. године и траје до данас. Седма пандемија није почела у Индији, већ у Индонезији, брзо се проширила на Филипине, Кину, земље Индокине, а затим и на друге земље Азије, Африке и Европе. Посебности ове пандемије укључују чињеницу да ју је, прво, изазвала посебна варијанта вибриона колере - V. cholerae eltor, која до 1961. године није била ни званично призната као узрочник колере; друго, по трајању је надмашила све претходне пандемије; треће, јавила се у два таласа, од којих је први трајао до 1990. године, а други је почео 1991. године и обухватио је многе земље Јужне и Северне Америке, укључујући Сједињене Америчке Државе, које нису виделе епидемију колере од 1866. године. Од 1961. до 1996. године од колере је оболело 3.943.239 људи у 146 земаља.
Узрочника колере, Vibrio cholerae, открио је Р. Кох 1883. године током пете пандемије, али је вибрион први пут открио у фецесу пацијената са дијарејом 1854. године Ф. Пачини.
V. cholerae припада породици Vibrionaceae, која обухвата неколико родова (Vibrio, Aeromonas, Plesiomonas, Photobacterium). Род Vibrio од 1985. године има више од 25 врста, од којих су за људе најважније V. cholerae, V. parahaemolyticus, V. alginolyticus, V. vulnificus и V. fluvialis.
Кључне карактеристике рода Vibrio
Кратки, неспоро- и капсулообразни, закривљени или прави грам-негативни штапићи, пречника 0,5 µм и дужине 1,5-3,0 µм, покретни (V. cholerae је монотрихозан, неке врсте имају два или више поларних флагела); добро и брзо расту на редовним медијумима, хемоорганотрофи су и ферментишу угљене хидрате да би произвели киселину без гаса (глукоза се ферментише путем Ембден-Мајерхофовог пута). Оксидаза-позитивни, формирају индол, редукују нитрате у нитрите (V. cholerae даје позитивну нитрозоиндолну реакцију), разлажу желатин, често дају позитивну Вогес-Проскауерову реакцију (тј. формирају ацетилметилкарбинол), немају уреазу, не формирају H2S, имају лизин и орнитин декарбоксилазе, али немају аргинин дихидролазу. Карактеристична особина рода Vibrio је осетљивост већине бактеријских сојева на лек 0/129 (2,4-диамино-6,7-диазопропилптеридин), док су представници породица Pseudomonadaceae и Enterobacteriaceae отпорни на овај лек. Вибриони су аероби и факултативни анаероби, температурни оптимум за раст је 18-37 C, pH 8,6-9,0 (расту у pH опсегу од 6,0-9,6), неке врсте (халофили) не расту у одсуству NaCl. Садржај G + C у ДНК је 40-50 mol % (за V. cholerae око 47 mol %). Биохемијски тестови се користе за разликовање унутар породице Vibrionaceae од морфолошки сличних родова Aeromonas и Plesiomonas, као и за разликовање од породице Enterobacteriaceae.
Колерни вибрион се разликује од породице Pseudomonadaceae по томе што ферментише глукозу само путем Ембден-Мајерхофовог пута (без учешћа О2), док први конзумирају глукозу само у присуству О2. Ова разлика између њих се лако открива на Хју-Лајфсоновој подлози. Подлога садржи хранљиви агар, глукозу и индикатор. Сетва се врши у две колоне са Хју-Лајфсоновом подлогом, од којих је једна напуњена вазелином (за стварање анаеробних услова). У случају раста колерног вибриона, боја подлоге се мења у обе епрувете, у случају раста псеудомонада - само у епрувети без вазелина (аеробни услови раста).
Колерни вибрион је веома незахтеван за хранљиве подлоге. Добро и брзо се размножава у 1% алкалној (pH 8,6-9,0) пептонској води (PV) која садржи 0,5-1,0% NaCl, престижући раст других бактерија. Да би се сузбио раст Протеуса, препоручује се додавање калијум телурита (у коначном разблажењу 1:100.000) у 1% PV. 1% PV је најбоља подлога за обогаћивање колерног вибриона. Током раста, након 6-8 сати формира нежан, растресит, сивкаст филм на површини PV, који се лако уништава када се протресе и пада на дно у облику пахуљица, PV постаје умерено замућен. За изолацију колерног вибриона предложене су различите селективне подлоге: алкални агар, агар са жучном сољу, алкални албуминат, алкални агар са крвљу, лактоза-сахароза и друге подлоге. Најбољи је TCBS (тиосулфат цитрат-бромотимол сахароза агар) медијум и његове модификације. Међутим, најчешће се користи алкални MPA, на коме колерни вибрион формира глатке, стакласто-провидне, плавкасто обојене, дисколике колоније вискозне конзистенције.
Када се посеје ињекцијом у колону желатина, вибрион, након 2 дана на температури од 22-23 C, изазива ликвефакцију са површине у облику мехурића, затим левкастог облика и коначно, слој по слој.
У млеку се вибрион брзо размножава, узрокујући коагулацију након 24-48 сати, а затим долази до пептонизације млека и након 3-4 дана вибрион умире због промене pH вредности млека на киселу страну.
Б. Хајберг је, на основу њихове способности ферментације манозе, сахарозе и арабинозе, поделио све вибрионе (холере и сличне колери) у неколико група, чији број сада износи 8.
Вибрио колере припада првој групи Хајберга.
Вибриони слични по морфолошким, културолошким и биохемијским карактеристикама холерном вибриону називани су и називају се различито: парахолера, холерни вибриони, NAG-вибриони (неаглутинирајући вибриони); вибриони који не припадају О1 групи. Презиме најтачније наглашава њихову везу са холерним вибрионом. Како су утврдили А. Гарднер и К. Венкат-Раман, холера и холерни вибриони имају заједнички Х-антиген, али се разликују по О-антигенима. Према О-антигену, холера и холерни вибриони су тренутно подељени у 139 О-серогрупа, али се њихов број стално шири. Холерни вибрион припада О1 групи. Има заједнички А-антиген и два типски специфична антигена - Б и Ц, помоћу којих се разликују три серотипа V. cholerae - Огава серотип (AB), Инаба серотип (AC) и Хикошима серотип (ABC). Вибрион колере у фази дисоцијације има OR-антиген. У том смислу, за идентификацију V. cholerae користе се О-серум, OR-серум и типски специфични серуми Инаба и Огава.
Године 1992-1993, у Бангладешу, Индији, Кини, Малезији и другим земљама почела је велика епидемија колере, чији је узрочник био нови, раније непознати серовар врсте Vibrio cholerae. Разликује се од V. cholerae O1 по антигеним карактеристикама: има антиген 0139 и полисахаридну капсулу и не аглутинише га ниједан други О-серум. Сва његова остала морфолошка и биолошка својства, укључујући способност изазивања колере, тј. синтезе егзотоксин-холерогена, показала су се сличним својствима V. cholerae O1. Сходно томе, нови узрочник колере, V. cholerae 0139, очигледно је настао као резултат мутације која је променила О-антиген. Назван је V. cholerae 0139 бенгал.
Питање односа такозваних вибриона сличних колери са V. cholerae дуго је било нејасно. Међутим, поређење V. cholerae и вибриона сличних колери (NAG-вибриона) по више од 70 карактеристика открило је њихову сличност од 90%, а степен ДНК хомологије V. cholerae и проучаваних NAG-вибриона је 70-100%. Стога се вибриони слични колери комбинују у једну врсту са вибрионом колере, од којег се разликују углавном по својим О-антигенима, у вези са чиме се називају вибриони не-01-групе - V. cholerae non-01.
Врсте V. cholerae су подељене на 4 биотипа: V. cholerae, V. eltor, V. proteus и V. albensis. Природа вибриона Ел Тор је предмет расправе већ дуги низ година. Овај вибрион је изоловао Ф. Готшлих 1906. године у карантинској станици Ел Тор из тела ходочасника који је умро од дизентерије. Ф. Готшлих је изоловао неколико таквих сојева. Они се нису разликовали од вибриона колере по свим својим својствима и били су аглутинисани О-серумом колере. Међутим, пошто у то време није било колере међу ходочасницима, а дугорочно ношење вибриона колере сматрало се мало вероватним, питање могуће етиолошке улоге V. eltor у колери остало је контроверзно дуго времена. Поред тога, вибрион Ел Тор, за разлику од V. cholerae, имао је хемолитички ефекат. Међутим, 1937. године, овај вибрион је изазвао велику и тешку епидемију колере на острву Сулавеси (Индонезија) са стопом морталитета преко 60%. Коначно, 1961. године, постао је виновник 7. пандемије, а 1962. године коначно је решено питање његове колерне природе. Разлике између V. cholerae и V. eltor тичу се само неких карактеристика. По свим осталим својствима, V. eltor се суштински не разликује од V. cholerae. Поред тога, сада је утврђено да биотип V. proteus (V.finklerpriori) обухвата целу групу вибриона, осим 01 групе (а сада 0139), раније назване NAG вибриони. Биотип V. albensis је изолован из реке Елбе и има способност фосфоресцирања, али пошто ју је изгубио, не разликује се од V. proteus. На основу ових података, врста Vibrio cholerae је тренутно подељена на 4 биотипа: V. cholerae 01 cholerae, V. cholerae eltor, V. cholerae 0139 bengal и V. cholerae non 01. Прва три припадају двама сероварима 01 и 0139. Последњи биовар обухвата претходне биотипове V. proteus и V. albensis и представљен је многим другим сероварима вибриона који нису аглутинисани серумима 01 и 0139, односно NAG вибрионима.
Фактори патогености колерног вибриона
 [ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ]
[ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ]
Хемотаксија вибриона колере
Уз помоћ ових својстава, вибрион интерагује са епителним ћелијама. Код мутаната колерног вибриона (који су изгубили способност хемотаксије), вируленција је значајно смањена, код мутаната Mob (који су изгубили покретљивост) она или потпуно нестаје или се нагло смањује.
Фактори адхезије и колонизације помоћу којих се вибрион причвршћује за микровиле и колонизује слузокожу танког црева. Фактори адхезије укључују муциназу, растворљиви хемаглутинин/протеазу, неураминидазу итд. Они подстичу адхезију и колонизацију уништавајући супстанце које су део слузи. Растворљиви хемаглутинин/протеаза подстиче одвајање вибриона од рецептора епителних ћелија и њихов излазак из црева у спољашњу средину, осигуравајући њихово епидемијско ширење. Неураминидаза јача везу између холерагена и епителних ћелија и олакшава продор токсина у ћелије, што повећава тежину дијареје.
Холерни токсин је холераген.
Такозвани нови токсини који су способни да изазову дијареју, али немају генетску или имунолошку везу са холерагеном.
Дермонеуротски и хеморагични фактори. Природа ових токсичних фактора и њихова улога у патогенези колере нису довољно проучене.
[ 13 ], [ 14 ], [ 15 ], [ 16 ], [ 17 ], [ 18 ]
Ендотоксини вибриона колере
Липополисахариди V. cholerae имају јако ендотоксично својство и изазивају општу интоксикацију организма.
Главни од наведених фактора патогености холерног вибриона је егзотоксин холераген (CTX AB), који одређује патогенезу ове болести. Молекул холере се састоји од два фрагмента - А и Б. Фрагмент А се састоји од два пептида - А1 и А2, има специфично својство холерног токсина и даје му квалитете суперантигена. Фрагмент Б се састоји од 5 идентичних подјединица. Обавља две функције: 1) препознаје рецептор (моносијалоганглиозид) ентероцита и везује се за њега; 2) формира интрамембрански хидрофобни канал за пролаз подјединице А. Пептид А2 служи за везивање фрагмената А и Б. Стварну токсичну функцију обавља пептид Ај (АДП-рибозилтрансфераза). Он интерагује са НАД-ом, узрокујући његову хидролизу; Добијена АДП-рибоза се везује за регулаторну подјединицу аденилат циклазе. То доводи до инхибиције хидролизе ГТП-а. Добијени комплекс ГТП + аденилат циклаза изазива хидролизу АТП-а са формирањем цАМП-а. (Други пут за акумулацију цАМП-а је супресија ензима који хидролизује цАМП до 5-АМП помоћу холерагена). Манифестација функције гена ctxAB који кодира синтезу егзотоксина зависи од функције низа других гена патогености, посебно гена tcp (који кодирају синтезу адхезионих пилула контролисаних токсином - TCAP), регулаторних гена toxR, toxS и toxT, гена hap (растворљиви хемаглутинин/протеаза) и неураминидазе (неураминидаза). Стога је генетска контрола патогености V. cholerae сложена.
Како се испоставило, постоје два острва патогености у хромозому V. cholerae. Једно од њих је геном филаментозног умерено конвертујућег фага CTXφ, а друго је геном такође филаментозног умерено конвертујућег фага VPIcp. Свако од ових острва патогености садржи касете гена наведених у профази, који одређују патогеност патогена колере. Профаг CTXφ носи гене CTX, гене нових токсина zot и ace, ген ser (синтеза адхезина) и ген ortU (синтеза производа са непознатом функцијом). Ова касета такође укључује ген nei и регион фага RS2, који кодира репликацију и интеграцију профага у хромозоме. Гени zot, ace и ortU су неопходни за формирање фагних вириона када је профаг искључен из хромозома патогена.
Профаг VPIcp носи гене tcp (који кодирају производњу пила (TCPA протеин)), toxT, toxR, act гене (додатни фактор колонизације, гени мобилности (интегразе и транспозазе)). Транскрипцију гена вируленције регулишу три регулаторна гена: toxR, toxS и toxT. Ови гени координирано, на нивоу транскрипције, мењају активност више од 20 гена вируленције, укључујући ctxAB, tcp и друге гене. Главни регулаторни ген је ген toxR. Његово оштећење или одсуство доводи до авируленције или до смањења производње колерног токсина CTX и TCPA за више од 100 пута. Могуће је да је на овај начин координирана експресија гена вируленције регулисана у острвима патогености које формирају умерени конвертујући фаги и код других бактеријских врста. Утврђено је да је још један профаг K139 присутан у хромозому V. cholerae eltor, али је његов геном мало проучен.
Ген hap је локализован на хромозому. Дакле, вируленцију (патогеност) и епидемијски капацитет V. cholerae одређују 4 гена: ctxAB, tcp, toxR и hap.
Различите методе се могу користити за откривање способности V. cholerae да производи холераген.
Биолошки тест на зечевима. Када се вибриони колере убризгају интрамускуларно зечевима који сисају (старости не више од 2 недеље), они развијају типичан синдром колере: дијареју, дехидрацију и смрт зеца.
Директна детекција холерогена помоћу PCR, IFM или реакције пасивне имуне хемолизе (холероген се везује за Gmj еритроцита, а они се лизирају након додавања антитоксичних антитела и комплемента). Међутим, само откривање способности производње токсина није довољно да се утврди епидемијска опасност таквих сојева. За то је неопходно детектовати присуство hap гена, стога је најбољи и најпоузданији начин за разликовање токсигених и епидемијских сојева холерних вибриона серогрупа 01 и 0139 PCR коришћењем специфичних прајмера за детекцију свих 4 гена патогености: ctxAB, tcp, toxR и hap.
Способност V. cholerae, осим серогрупа 01 или 0139, да изазове спорадичне или кластер дијарејне болести код људи може бити последица присуства ентеротоксина типа LT или ST, који стимулишу аденилатни односно гванилат циклазни систем, или присуства само гена ctxAB, али не и гена hap.
Током седме пандемије, изоловани су сојеви V. cholerae са различитим степеном вируленције: холерогени (вирулентни), слабо холерогени (ниске вируленције) и нехолерогени (невирулентни). Нехолерогени V. cholerae, по правилу, показују хемолитичку активност, не лизирају се помоћу дијагностичког фага колере HDF(5) и не изазивају болести код људи.
За фаготипизацију V. cholerae 01 (укључујући El Tor) С. Мукерџи је предложио скупове фага, који су потом у Русији допуњени другим фагима. Скуп таквих фага (1-7) омогућава разликовање типова фага међу V. cholerae 0116. За идентификацију токсигених и нетоксигених V. cholerae El Tor, уместо HDF-3, HDF-4 и HDF-5, у Русији се сада предлажу фаги CTX* (лизујући токсигене El Tor вибрионе) и CTX" (лизујући нетоксигене El Tor вибрионе).
Отпорност узрочника колере
Вибриони колере добро преживљавају на ниским температурама; остају одрживи у леду до 1 месец; у морској води - до 47 дана, у речној води - од 3-5 дана до неколико недеља, у кувани минералној води преживљавају више од 1 године, у земљишту - од 8 дана до 3 месеца, у свежем фецесу - до 3 дана, на куваним производима (пиринач, резанци, месо, каша итд.) преживљавају 2-5 дана, на сировом поврћу - 2-4 дана, на воћу - 1-2 дана, у млеку и млечним производима - 5 дана; када се чувају на хладном, период преживљавања се повећава за 1-3 дана; на ланеном платну контаминираном фецесом преживљавају до 2 дана, а на влажном материјалу - недељу дана. Вибриони колере умиру у року од 5 минута на температури од 80 °C, а тренутно на 100 °C; веома су осетљиви на киселине; умиру у року од 5-15 минута под утицајем хлорамина и других дезинфекционих средстава. Осетљиве су на сушење и директну сунчеву светлост, али добро и дуго преживљавају и чак се размножавају у отвореним воденим површинама и отпадним водама богатим органским материјама, са алкалним pH и температуром изнад 10-12 °C. Веома су осетљиве на хлор: доза активног хлора од 0,3-0,4 мг/л воде за 30 минута изазива поуздану дезинфекцију од вибриона колере.
Вибриони патогени за људе који не припадају врсти Vibrio Cholerae
Род Vibrio обухвата више од 25 врста, од којих је, поред V. cholerae, најмање осам способно да изазове болести код људи: V. parahaemolyticus, V. alginolyticus, V. vulnificus, V. fluvialis, V. fumissii, V. mimicus, V. damsela и V. hollisae. Сви ови вибриони насељавају мора и заливе. Инфекција се јавља или пливањем или конзумирањем морских плодова. Утврђено је да колерни и не-холерни вибриони могу изазвати не само гастроентеритис, већ и инфекције рана. Ова способност је пронађена код V. cholerae 01 и не-01 групе, V. parahaemolyticus, V. alginolyticus, V. mimicus, V. damsela и V. vulnificus. Они изазивају инфламаторне процесе у меким ткивима када их оштети љуштура морских животиња или када су у директном контакту са зараженом морском водом.
Од наведених патогених не-холерних вибриона, највећи практични интерес су V. parahaemolyticus, V. alginolyticus, V. vulnificus и V. fluvialis.
V. parahaemolyticus - парахемолитички вибрион - први пут је изолован у Јапану 1950. године током велике епидемије тровања храном изазване конзумирањем полусушених сардина (морталитет је био 7,5%). Узрочник је припадао роду Vibrio, а открио га је Р. Саказаки 1963. године. Он је проучаване сојеве поделио на 2 врсте: V. parahaemolyticus и V. alginolyticus. Обе врсте се налазе у приобалној морској води и код њених становника, они су халофили (грч. hals - со); за разлику од обичних вибриона, халофилни не расту на медијумима без NaCl и добро се размножавају при високим концентрацијама истог. Припадност врстама халофилних вибриона одређена је њиховом способношћу да ферментишу сахарозу, формирају ацетилметилкарбинол и размножавају се у ПВ са 10% NaCl. Све ове особине су својствене врсти V. alginolyticus, али су одсутне код V. parahaemolyticus.
Парахемолитички вибрион има три врсте антигена: термолабилне флагеларне Х-антигене, термостабилне О-антигене који се не уништавају загревањем на 120 °C током 2 сата и површинске К-антигене који се уништавају загревањем. Свеже изоловане културе V. parahaemolyticus имају добро дефинисане К-антигене који штите живе вибрионе од аглутинације хомологним О-серумима. Х-антигени су исти за све сојеве, али се Х-антигени монотрихуса разликују од Х-антигена перитрихуса. Према О-антигену, V. parahaemolyticus је подељен на 14 серогрупа. Унутар серогрупа, вибриони су подељени на серотипове према К-антигенима, чији је укупан број 61. Антигенска шема V. parahaemolyticus је развијена само за његове сојеве изоловане од људи.
Патогеност V. parahaemolyticus повезана је са његовом способношћу да синтетише хемолизин, који има ентеротоксична својства. Потоња се детектује Канагава методом. Њена суштина лежи у чињеници да V. parahaemolyticus, патоген за људе, изазива јасну хемолизу на крвном агару који садржи 7% NaCl. На крвном агару који садржи мање од 5% NaCl, хемолизу изазивају многи сојеви V. parahaemolyticus, а на крвном агару са 7% NaCl - само сојеви са ентеропатогеним својствима. Парахемолитички вибрион се налази на обалама Јапанског, Каспијског, Црног и других мора. Изазива токсичне инфекције које се преносе храном и болести сличне дизентерији. Инфекција се јавља конзумирањем сирових или полусирових морских плодова заражених V-parahaemolyticus (морске рибе, остриге, ракови итд.).
Међу осам горе поменутих вибриона који нису колера, најпатогенији за људе је V. vulnificus, који је први пут описан 1976. године као Beneckea vulnificus, а затим рекласификован као Vibrio vulnificus 1980. године. Често се налази у морској води и њеним становницима и изазива разне људске болести. Сојеви V. vulnificus морског и клиничког порекла се не разликују једни од других ни фенотипски ни генетски.
Инфекције рана изазване V. vulnificus брзо напредују и доводе до стварања тумора са накнадном некрозом ткива, праћено грозницом, дрхтавицом, понекад јаким болом, а у неким случајевима захтевају ампутацију.
Утврђено је да V. vulnificus производи егзотоксин. Експерименти на животињама су показали да патоген изазива тешка локална оштећења са развојем едема и некрозе ткива, након чега следи смрт. Улога егзотоксина у патогенези болести се испитује.
Поред инфекција рана, V. vulnificus може изазвати упалу плућа код жртава утапања и ендометритис код жена након излагања морској води. Најтежи облик инфекције коју изазива V. vulnificus је примарна септикемија повезана са конзумирањем сирових острига (а могуће и других морских животиња). Ова болест се развија веома брзо: пацијент развија малаксалост, грозницу, језу и прострацију, затим тешку хипотензију, што је главни узрок смрти (стопа морталитета је око 50%).
В. флувијалис је први пут описан као патоген гастроентеритиса 1981. године. Припада подгрупи не-холерних патогених вибриона који имају аргинин дихидролазу, али не и неторнитин и лизин декарбоксилазе (В. флувијалис, В. furnissii, В. дамсела, тј. фенотипски сличне Аеромонасу). В. флувијалис је чест узрочник гастроентеритиса, који је праћен јаким повраћањем, дијарејом, боловима у стомаку, грозницом и тешком или умереном дехидрацијом. Главни патогени фактор је ентеротоксин.
Епидемиологија колере
Главни извор инфекције је само особа - болесник колере или вибрионоша, као и вода контаминирана њима. Ниједна животиња у природи не оболева од колере. Пут инфекције је феко-орални. Путеви инфекције: а) главни - кроз воду која се користи за пиће, купање и кућне потребе; б) контактно-кућни и ц) кроз храну. Све веће епидемије и пандемије колере биле су повезане са водом. Холерни вибриони имају такве адаптивне механизме који обезбеђују постојање њихових популација како у људском телу, тако и у одређеним екосистемима отворених водених тела. Тешка дијареја, коју изазива холерни вибрион, доводи до чишћења црева од конкурентских бактерија и доприноси широком ширењу патогена у животној средини, пре свега у отпадним водама и у отвореним воденим телама где се оне бацају. Особа оболела од колере излучује узрочника у огромним количинама - од 100 милиона до 1 милијарде на 1 мл фецеса, вибрионосац излучује 100-100.000 вибриона у 1 мл, инфективна доза је око 1 милион вибриона. Трајање излучивања холерног вибриона код здравих носилаца је од 7 до 42 дана и 7-10 дана код оних који су се опоравили. Дуже излучивање је изузетно ретко.
Посебност колере је да након ње, по правилу, нема дуготрајног ношења и не формирају се стабилна ендемска жаришта. Међутим, као што је већ горе назначено, због загађења отворених водених тела отпадним водама које садрже велике количине органских супстанци, детерџената и кухињске соли, лети колерни вибрион не само да дуго преживљава у њима, већ се чак и размножава.
Од великог епидемиолошког значаја је чињеница да вибриони колере 01 групе, како нетоксигени тако и токсигени, могу дуго времена да перзистирају у различитим воденим екосистемима као некултивисани облици. Коришћењем полимеразне ланчане реакције, vct гени некултивисаних облика V. chokrae су откривени у различитим воденим тијелима на бројним ендемским територијама ЗНД током негативних бактериолошких студија.
Ендемско жариште вибриона колере Ел Тор је Индонезија, појава овог виновника седме пандемије одатле се сматра повезаном са ширењем економских веза Индонезије са спољним светом након стицања независности, а на трајање и муњевит развој пандемије, посебно њеног другог таласа, пресудно је утицао недостатак имунитета на колеру и разни друштвени превирања у земљама Азије, Африке и Америке.
У случају колере, предузима се низ антиепидемијских мера, међу којима су водеће и одлучујуће активно, благовремено откривање и изолација (хоспитализација, лечење) пацијената у акутним и атипичним облицима и здравих вибрионосаца; предузимају се мере за спречавање могућих путева инфекције; посебна пажња се посвећује водоснабдевању (хлорисање воде за пиће), поштовању санитарно-хигијенских услова у прехрамбеним предузећима, у дечјим установама, јавним местима; спроводи се строга контрола, укључујући бактериолошку, над отвореним воденим површинама, спроводи се имунизација становништва итд.
Симптоми колере
Период инкубације за колеру варира од неколико сати до 6 дана, најчешће 2-3 дана. Након уласка у лумен танког црева, холерни вибриони, због своје покретљивости и хемотаксије до слузокоже, усмеравају се ка слузи. Да би продрли кроз њу, вибриони производе низ ензима: неураминидазу, муциназу, протеазе, лецитиназу, који уништавају супстанце садржане у слузи и олакшавају кретање вибриона до епителних ћелија. Адхезијом, вибриони се везују за гликокаликс епитела и, губећи покретљивост, почињу интензивно да се размножавају, колонизујући микровиле танког црева (видети уметак у боји, сл. 101.2), и истовремено производе велику количину егзотоксина-холерогена. Молекули холерагена везују се за моносијалоганглиозид Гни! И продиру кроз ћелијску мембрану, где активирају аденилат циклазни систем, а акумулирани цАМП изазива хиперсекрецију течности, катјона и ањона Na, HCO, Kl, Cl из ентероцита, што доводи до колерне дијареје, дехидрације и десалинизације организма. Постоје три врсте болести:
- насилна, тешка дехидрирајућа дијарејска болест која доводи до смрти пацијента у року од неколико сати;
- мање тежак ток или дијареја без дехидрације;
- асимптоматски ток болести (ношење вибриона).
У тешким случајевима колере, пацијенти развијају дијареју, учесталост столице се повећава, фецес постаје обилнији, постаје воденаст, губи мирис фекалије и изгледа као пиринчана чорба (мутна течност са остацима слузи и епителним ћелијама које плутају у њој). Затим долази до исцрпљујућег повраћања, прво цревног садржаја, а затим повраћање поприма изглед пиринчане чорбе. Температура пацијента пада испод нормале, кожа постаје плавкаста, наборана и хладна - холера алгид. Као резултат дехидрације, крв се згушњава, развија се цијаноза, гладовање кисеоником, функција бубрега нагло пати, јављају се конвулзије, пацијент губи свест и наступа смрт. Стопа морталитета од колере током седме пандемије варирала је од 1,5% у развијеним земљама до 50% у земљама у развоју.
Пост-инфективни имунитет је јак, дуготрајан, а рекурентне болести су ретке. Имунитет је антитоксичан и антимикробни, узрокован антителима (антитексини дуже трају од антимикробних антитела), ћелијама имуне меморије и фагоцитима.
Лабораторијска дијагностика колере
Главна и одлучујућа метода дијагностиковања колере је бактериолошка. Материјал за преглед од пацијента је фецес и повраћање; фецес се испитује на носиоштво вибриона; од особа које су умрле од колере узима се лигирани део танког црева и жучне кесе; од предмета спољашње средине најчешће се испитује вода из отворених резервоара и отпадне воде.
Приликом спровођења бактериолошке студије морају бити испуњена следећа три услова:
- што је брже могуће узети материјал од пацијента (колера вибрио преживљава у фецесу кратак временски период);
- посуда у којој се материјал узима не сме бити дезинфикована хемикалијама и не сме садржати њихове трагове, јер је колерни вибрион веома осетљив на њих;
- елиминисати могућност контаминације и инфекције других.
Култура се изолује према следећој шеми: сетва на ПВ, истовремено на алкалну МПА или било коју селективну подлогу (најбоље је ТЦБС). Након 6 сати, филм формиран на ПВ се испитује, и ако је потребно, врши се пренос на другу ПВ (брзина сетве холерног вибриона у овом случају се повећава за 10%). Из ПВ се врши пренос у алкалну МПА. Сумњиве колоније (стакласто-провидне) се преносе да би се добила чиста култура, која се идентификује морфолошким, културним, биохемијским својствима, покретљивошћу и коначно типизира коришћењем дијагностичких аглутинирајућих серума О-, ОР-, Инаба и Огава и фага (ХДФ). Предложене су различите опције за убрзану дијагностику, од којих је најбоља луминесцентно-серолошка метода. Она омогућава детекцију холерног вибриона директно у испитиваном материјалу (или након претходне култивације у две епрувете са 1% ПВ, од којих се једној дода холерни фаг) у року од 1,5-2 сата. За убрзано откривање вибриона колере, Нижњеновгородски ИЕМ је предложио сет папирних индикаторских дискова који се састоји од 13 биохемијских тестова (оксидаза, индол, уреаза, лактоза, глукоза, сахароза, маноза, арабиноза, манитол, инозитол, аргинин, орнитин, лизин), који омогућавају разликовање представника рода Vibrio од родова Aeromonas, Plesiomonas, Pseudomonas, Comamonas и из породице Enterobacteriaceae. За брзо откривање вибриона колере у фецесу и у објектима животне средине, може се користити РПГА са дијагностичким антителима. Да би се открили некултивисани облици вибриона колере у објектима животне средине, користи се само метода ланчане реакције полимеразе.
У случајевима када су изоловане колере V. које нису из Ol-групе, треба их типизирати коришћењем одговарајућих аглутинирајућих серума других серогрупа. Изолација колера V. које нису из Ol-групе од пацијента са дијарејом (укључујући дијареју сличну колери) захтева исте антиепидемијске мере као у случају изолације колера V. Ol-групе. Ако је потребно, присуство гена патогености ctxAB, tcp, toxR и hap се у таквим вибрионима одређује помоћу PCR.
Серолошка дијагностика колере је помоћне природе. У ту сврху може се користити реакција аглутинације, али је боље одредити титар вибриоцидних антитела или антитоксина (антитела на колеру се одређују ензимским имунотестом или имунофлуоресцентним методама).
Лабораторијска дијагностика непатогених вибриона који нису изазвани колером
Главна метода дијагностиковања болести изазваних не-холерним патогеним вибрионима је бактериолошка коришћењем селективних медија као што су TCBS, MacConkey итд. Припадност изоловане културе роду Vibrio се одређује на основу кључних карактеристика бактерија овог рода.
Лечење колере
Лечење оболелих од колере треба првенствено да се састоји од рехидратације и обнављања нормалног метаболизма воде и соли. У ту сврху препоручује се употреба физиолошких раствора, на пример, следећег састава: NaCl - 3,5; NaHC03 - 2,5; KCl - 1,5 и глукоза - 20,0 г на 1 литар воде. Такав патогенетски утемељен третман у комбинацији са рационалном антибиотском терапијом омогућава смањење стопе морталитета код колере на 1% или мање.
Специфична превенција колере
За стварање вештачког имунитета предложена је вакцина против колере, укључујући ону направљену од убијених сојева Инаба и Огава; токсоид колере за поткожну употребу и ентерална хемијска бивалентна вакцина која се састоји од анатоксина и соматских антигена серотипова Инаба и Огава, пошто се не формира унакрсни имунитет. Међутим, трајање поствакциналног имунитета није дуже од 6-8 месеци, па се вакцинације спроводе само према епидемиолошким индикацијама. Антибиотска профилакса се добро показала у жариштима колере, посебно тетрациклин, на који је вибрион колере веома осетљив. У исту сврху могу се користити и други антибиотици ефикасни против V. cholerae.